
MEMBRANA
A célula eucariótica constitui um espaço fortemente compartimentado. Desta compartimentação decorre implicitamente a especialização de certas regiões ou estruturas para o desempenho de funções determinadas.
Com efeito, as numerosas reacções bioquímicas que ocorrem simultaneamente no espaço celular encontram-se fisicamente confinadas em compartimentos delimitados por estruturas membranares. São deste modo, por um lado, evitadas interferências espontâneas entre reacções antagónicas, as quais seriam indutoras de desordem. Por outro, a existência de proteínas, com funções enzimáticas, inseridas na membrana, permite que certas reacções possam ocorrer junto da superfície destas, de forma ordenada, sequencial.
De acordo com o conceito trifásico, todo o funcionamento da célula assenta no intercâmbio entre as duas fases e, destas, com o exterior. O conhecimento da composição química, da arquitectura molecular, das propriedades e das funções das membranas celulares é por conseguinte, essencial à compreensão dos fenómenos subjacentes à vida da célula. A sua arquitectura molecular sendo universal, não exclui contudo diferenças sectoriais ao nível da composição química e da própria espessura, relacionadas com as funções que exerce.
Composição química da membrana
As membranas constituintes das células procarióticas como eucarióticas, são lâminas finas e deformáveis, mas mecanicamente resistentes; são todas estruturadas de acordo com o mesmo modelo de arquitectura molecular, ainda que possam apresentar diferentes espessuras (6 –10 nm), razão pela qual J. David Robertson as designou por membrana unitária.
A composição química das membranas oscila em torno dos valores médios de 60% de proteínas globulares e 40% de lípidos. Associados a estes componentes maioritários, identificam-se ainda glúcidos, quase sempre em quantidades muito menores e associados às proteínas e aos lípidos, constituindo glicoproteínas e glicolípidos.
Entre as proteínas constituintes da membrana, cerca de 80% são enzimas. Os lípidos das membranas são moléculas longas e anfipáticas: possuem duas extremidades com propriedades de solubilidade, diferentes. Enquanto que uma das extremidades é hidrofílica (polar) e portanto solúvel em meio aquoso, a outra é hidrófoba (apolar), consequentemente insolúvel em meio aquoso mas com afinidade para outros lípidos. Entre os lípidos mais frequentes nas membranas celulares, distinguem-se os fosfolípidos, com uma representação de 70 a 90%. As membranas das células animais contêm colesterol, o que não acontece nas células vegetais, que possuem outros esterois. Como se verá adiante, quanto maior for a concentração de esterois, menos fluida será a membrana. As membranas das células procarióticas não contêm esterois, salvo raras excepções.
Arquitectura molecular
À medida que avança o conhecimento sobre a composição, a estrutura e as funções da membrana unitária, formulam-se modelos interpretativos dos dados conhecidos.
O primeiro modelo concebido, inicialmente por Gorder e Grendel, e depois desenvolvido por Danielli, Davson e outros, ficou conhecido por modelo em sandwich. Admitia que a membrana era formada por uma bicamada contínua de moléculas lipídicas, à qual se associavam proteínas, numa e noutra face. Este modelo de organização estrutural membranar deduzia-se a partir de dados físicos e químicos indirectos, nomeadamente na composição química da membrana e nas propriedades anfipáticas dos fosfolípidos. Com o desenvolvimento do microscópio electrónico de transmissão tornou-se possível visualizar directamente a estrutura da membrana, permitindo dar um importante passo em frente. A microscopia electrónica revelou uma estrutura tri-lamelar, consistindo em duas camadas electronodensas separadas por uma electronotranslúcida.
O arranjo dos lípidos em bicamada resulta directamente da sua natureza anfipática, acima definida. Como consequência disso, nas bicamadas lipídicas, as “cabeças” hidrofílicas das moléculas de lípidos ficam voltadas para fora, interactuando com os meios aquosos, enquanto que as “caudas” hidrófobas se dirigem umas para as outras, no interior da bicamada.
A dificuldade de coadunar o modelo com as propriedades de permeabilidade conhecidas, levou Danielli a admitir a existência de poros.
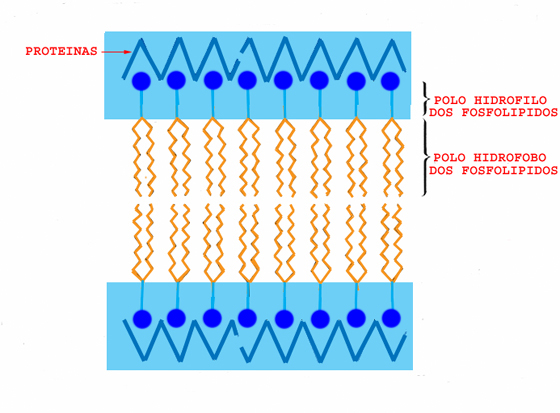
Modelo em sandwich (Davson e Danielli). A: corte transversal da membrana observado em microscopia
electrónica. B: esquema
Se a observação de cortes ultrafinos, em microscopia electrónica, permitiu formar um conceito geral da estrutura da membrana, a autêntica revolução da nossa compreensão da arquitectura membranar não surgiu senão com o advento de outra técnica de microscopia electrónica, a criofractura. Esta técnica permite observar, não o perfil, mas a superfície e, nomeadamente, a superfície interna, uma vez separados os dois folhetos lipídicos da membrana.
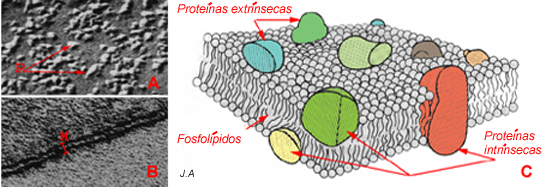
Modelo em “mosaico fluido” (Singer e Nicholson). A: imagem da superfície interna da
membrana, obtida por criofractura e observada em microscopia electrónica, mostrando as proteínas (P) . B: esquema.
De posse dos resultados disponibilizados pela nova técnica de microscopia electrónica, Singer, Nicholson e outros conceberam um novo modelo de arquitectura molecular, que ficou conhecido por modelo em mosaico fluido. Este recupera do anterior, a disposição dos fosfolípidos em dupla camada. Contudo, as proteínas, não só se dispõem de um e de outro lado, como também penetram na camada lipídica, podendo ocupar toda a espessura da membrana.
As proteínas, por seu turno, podem encontrar-se associadas à membrana essencialmente de duas formas distintas: ou se encontram “mergulhadas” na bicamada lipídica e designam-se por proteínas intrínsecas, ou encontram-se aderentes a uma das faces da membrana, designam-se então por proteínas extrínsecas .
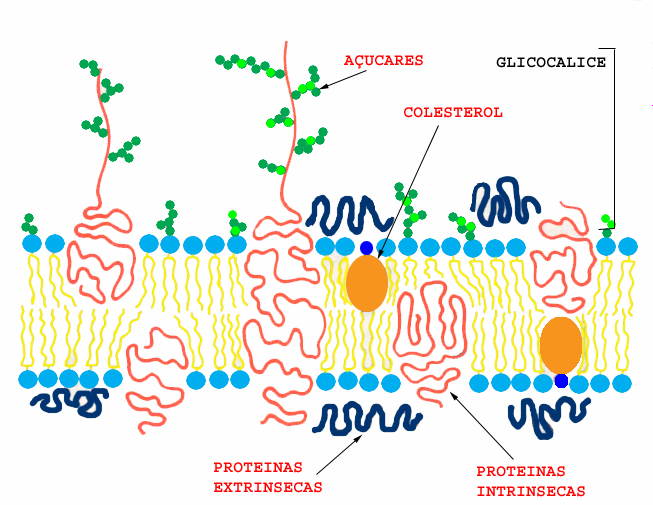
Em certos casos, as arborescências glucídicas dos glicolípidos e das glicoproteínas, em superfície da membrana plasmática, são muito abundantes e constituem um revestimento externo da célula, designado por glicocálice ou cell
Propriedades da membrana
As membranas gozam de um conjunto de propriedades físicas que determinam a sua participação não só na estruturação física da célula, como no próprio funcionamento. Destacam-se as seguintes propriedades:
a) Assimetria
As duas faces da membrana não possuem a mesma composição lipídica, glucídica e proteica. Em geral, os glúcidos encontram-se presentes na face externa. Também as cargas eléctricas se distribuem diferentemente, sendo a face citoplasmática, a que tem maior carga negativa, em geral.
b) Fluidez
A membrana é uma estrutura fluida, o que significa que os seus componentes não ocupam posições definidas e são susceptíveis de deslocações bidimensionais, de rotação ou de translação. Esta propriedade deve-se ao facto de, em geral, não se estabelecerem ligações fortes (covalentes) entre as diversas moléculas, mas, predominantemente, ligações lábeis (ligações de Van der Walls e pontes de hidrogénio). Para além dos movimentos referidos, também os fosfolípidos podem trocar de camada (flip-flop).
c) Permeabilidade diferenciada
A bicamada lipídica da membrana é impermeável aos iões, diferentemente permeável às moléculas consoante o respectivo peso molecular e lipossolubilidade, e francamente permeável à água e aos gases (azoto, oxigénio, dióxido de carbono). Aplica-se-lhe, ainda que de forma pouco rigorosa, o conceito de membrana semi-permeável.
d) Continuidade
Uma característica das membranas celulares reside no facto de elas nunca apresentarem bordos livres ou descontinuidades; em todas as formações, desde a membrana plasmática, que limita a célula, à mais pequena vesícula, não existem descontinuidades, e os espaços por ela delimitados, são sempre fechados.
A existência de poros nucleares explica-se pelo facto de o núcleo não ser delimitado por uma só membrana, mas por um invólucro duplo, constituído por duas membranas paralelas. Estas estão aliás em perfeita continuidade com as membranas do retículo endoplasmático, de tal forma que é lícito encarar o invólucro nuclear como uma parte especializada do próprio retículo endoplasmático.
e) Resistência à tracção
Apesar de os diversos componentes, nomeadamente os fosfolípidos constituintes da bicamada, estarem unidos por ligações fracas, o certo é que a integração dessas forças, em número extremamente elevado, confere à membrana uma determinada resistência à tracção, responsável pela manutenção da individualidade da célula. A indução da hemólise dos glóbulos vermelhos, através da sujeição destes a um meio hipotónico, põe em destaque o limiar da resistência da membrana.
Funções da membrana
À luz do conceito trifásico da célula eucariótica, cabe às membranas não só assegurar os contactos com o meio exterior, como desempenhar o papel de medianeiro (fase intermédia) entre as outras duas fases, a interna e a externa. Daí decorrem as principais funções da membrana:
a) Individualização da célula
O somatório das forças de Van der Walls que unem os fosfolípidos lado a lado, nas duas camadas, confere à membrana uma determinada resistência à tracção (ver propriedades da membrana), suficiente para assegurar a integridade física da célula, em condições normais e, consequentemente, a sua individualidade. Sabe-se contudo, que a membrana plasmática é complementada, nesta função, internamente, pelo citosqueleto, e externamente, por matrizes extracelulares (caso de algumas células animais) e pela parede celular (caso das células vegetais).
b) Transportes moleculares e iónicos
O relacionamento da célula com o meio exterior, bem como das duas fases, interna e externa, entre si, implica a permuta de substâncias: iões, moléculas pequenas, macromoléculas e mesmo partículas. Todas essas permutas se realizam através da membrana, com recurso a diversos mecanismos de transporte membranar, que garantem a selectividade do processo e o fluxo consentâneo com as necessidades da célula.
c) Recepção de informação
A adequação do funcionamento da célula às condições envolventes, sejam elas o meio exterior, sejam as que resultam de um contexto de organização pluricelular, implica a capacidade de recepção e de processamento de informação. Mais uma vez, compete à membrana, e em particular à membrana plasmática, assegurar a recepção da informação pertinente. Esta função é exercida, nomeadamente, em relação a informação consubstanciada em hormonas e feromonas, através de receptores proteicos incorporados na membrana.
d) Transmissão de informação
Algumas células, animais e vegetais, estabelecem, entre si, mecanismos de transmissão de informação, instalados ao nível da membrana. No caso concreto das células nervosas, a transmissão de informação constitui a função primordial para a qual se orienta todo o processo de especialização. A membrana assume, neste processo, funções determinantes, quer constituindo estruturas isoladoras (mielina), quer assegurando a propagação de um sinal eléctrico (potencial de acção).
e) Reconhecimento celular
Esta função é particularmente importante no caso das células que se organizam em sociedades pluricelulares (organismos), nas quais se estabelece, com base num património genético comum, uma diferenciação de funções. A garantia de que as células constituintes do organismo pertencem à mesma “família” implica, não só a existência de sinais exteriores identitários, como a capacidade de os reconhecer. Correlativamente, implica que certas células, dentro do quadro de partilha de funções, disponham de mecanismos específicos de eliminação os eventuais “intrusos”. O reconhecimento celular é pois uma função multifacetada, na qual a membrana intervém a diversos títulos.
f) Orientação vectorial de reacções
Muitos dos processos químicos dos sistemas biológicos implicam longas sequências de reacções coordenadas. Por exemplo, a substância A é convertida em B, a B é convertida em C, e assim sucessivamente até ao produto final, sendo que cada uma das reacções é catalisada por um enzima (E1, E2, E3, etc.).
A eficiência do processo depende da disponibilidade dos reagentes para intervirem no momento certo, o que poderá não acontecer se se encontrarem dispersos no citoplasma. Pelo contrário, se os diversos enzimas envolvidos no processo se dispuserem sequencialmente à superfície da membrana, o produto de uma reacção passa a estar fisicamente disponível para intervir na reacção seguinte. A sequência de reacções processar-se-á com maior eficiência.
Diversos são os exemplos de reacções vectoriais orientadas pela disposição dos enzimas à superfície da membrana (ver fosforilação oxidativa)
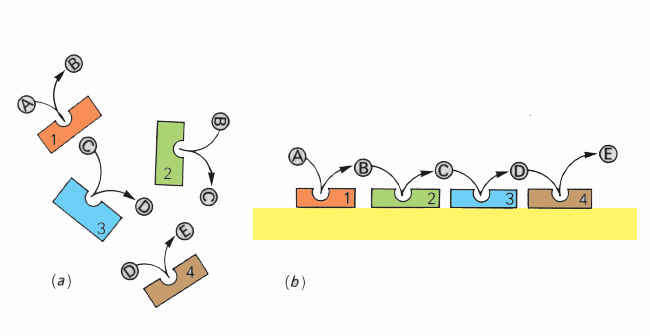
(a) Enzimas, reagentes e produtos dispersos no citoplasma
(b) Reacções orientadas pela disposição dos enzimas à superfície da membra
Especializações da membrana
Em regra, as células, quer se constituam como organismos unicelulares, quer se encontrem associadas formando organismos pluricelulares, apresentam especializações diversas, direccionadas a determinadas funções. Algumas dessas funções são estreitamente dependentes da membrana plasmática, como sejam a absorção de nutrientes ou a coesão intercelular quando se trata de formação de tecidos de revestimento (epitélios).
a) Aumento da superfície de trânsito molecular
A especialização na função de absorção de nutrientes ou, de uma forma geral, de transferência molecular, traduz-se, em geral, numa hipertrofia da membrana, que pode concretizar-se pela diferenciação de estruturas digitiformes, denominadas microvilosidades.
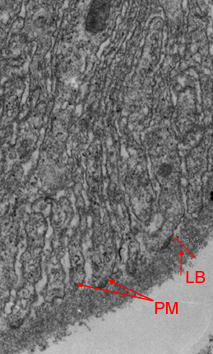
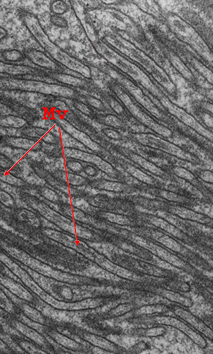
a) pregas membranares (P.M) recobertas por membrana basal (glicocálice) (LB); b) microvilosidades (MV)
As células epiteliais do intestino humano, por exemplo, possuem, na superfície apical, milhares de microvilosidades, que aumentam a área de absorção em cerca de 20 vezes. Noutros casos, o aumento da superfície de trânsito molecular resulta da formação de pregas membranares.
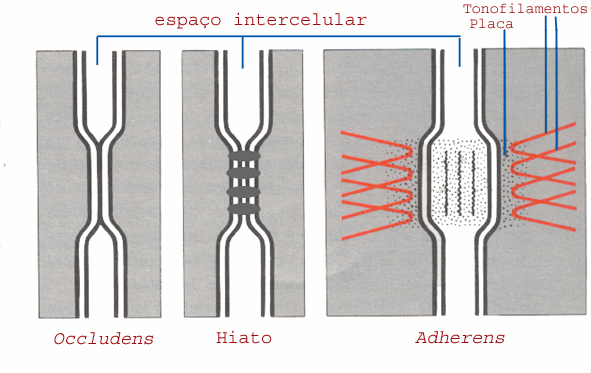
b) Coesão intercelular
Entre células adjacentes, constituintes de um tecido, subsiste um espaço intercelular de 10-15 nm. Contudo, em certos locais, formam-se zonas de aderência acrescida, designadas por junções. Distinguem-se, grosso modo, três classes de junções, consoante a função prioritária que exercem: impermeabilização, aderência, e comunicação.
Impermeabilização
Esta função é executada por junções apertadas ou zonulae ocludens. Destinam-se a tornar um tecido epitelial totalmente estanque. Consistem em bandas que rodeiam completamente as células e contactam com estruturas idênticas das células adjacentes. Ao seu nível, as membranas encostam-se, eliminando o espaço intermembranar.
Aderência
O reforço da aderência entre células adjacentes é realizado através das junções designadas por ou zonulae adherens (quando constituem bandas circundantes das células) ou fascia adherens, quando se limitam a pequenas zonas circulares. A este nível, o espaço intermembranar aumenta (15-20 nm) e é preenchido por um cimento glicoproteico. A membrana plasmática é mais espessa e dela partem tonofilamentos que penetram no citoplasma, oferecendo uma perspectiva de enraizamento. Incluem-se, nesta categoria, os desmossomas. Nestes últimos, o espaço intermembranar ainda é mais amplo, atingindo 30 nm, e as membranas das células vizinhas são acompanhadas, na face interna, por uma placa densa onde se prendem numerosos filamentos citoplasmáticos.
Comunicação
Nas células animais encontram-se junções em que o espaço intercelular se reduz a cerca de 3 nm e é, aparentemente, atravessado por septos. Estas junções são designadas por junções hiato ou gap junctions. O estudo ultra-estrutural destas junções revelou que os referidos septos são, na realidade, estruturas proteicas intrínsecas, formadas por seis proteínas idênticas, e designadas por conexões.
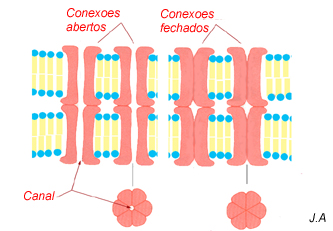
Os conexões de duas células adjacentes formam um canal contínuo que permite o trânsito de pequenas moléculas e de iões. Modificações estruturais das proteínas constituintes dos conexões, induzidas por alterações dos ambientes citoplasmáticos, como por exemplo o teor de Ca++, determinam que os canais estejam abertos ou fechados. As junções hiato são, pois, estruturas dinâmicas, e a sua participação em processos que implicam coordenação intercelular, como o peristaltismo intestinal, a contracção cardíaca, a embriogénese e a inibição de proliferação celular, por contacto entre as células, é conhecida.
Nos tecidos vegetais, a comunicação entre células adjacentes efectua-se através de plasmodesmos. Consistem em interrupções da parede celular preenchidas pelo citossol e atravessadas por um canal, o desmotúbulo, que garante a comunicação entre os retículos das duas células.
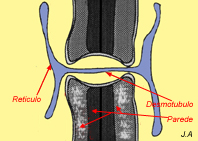
Plasmodesmos
Geralmente, os plasmodesmos formam-se durante a divisão celular. Apesar das suas dimensões relativamente grandes (20-40 nm), os plasmodesmos apresentam uma certa selectividade. Contudo, é através deles que certos vírus se propagam e infectam novas células.
______________________________________________________________________________________________________________
RETORNO