|
|
UNIVERSIDADE DE ÉVORA LABORATÓRIO DE BIOLOGIA MOLECULAR Engenharia Genética - Clonagem de DNAs em Bactérias (Módulo I) ACÇÃO FOCO - 1998 |
|
![[Universidade de Evora]](ue.gif)
|
ELECTROFORESE EM MACRO-MOLÉCULAS |
||
|
Ana Paula Vidigal |
Maria Clara Paes |
Maria Teresa Silva |
ELECTROFORESE DE MACROMOLÉCULAS
Diferentes tipos de electroforese
Aplicações práticas da electroforese
Trabalho sobre:
ELECTROFORESE DE MACROMOLÉCULAS
Chama-se electroforese ao transporte de partículas num campo eléctrico.
O processo é formalmente idêntico á sedimentação, já que em ambos os casos um campo, que é o gradiente de um potencial, produz o transporte das moléculas. No transporte electroforético, à força do campo opõe-se a resistência viscosa do meio, produzindo, quando se igualam, uma velocidade constante das partículas.
A electroforese aplica-se no campo da bioquímica na separação de compostos que possuem carga (aminoácidos, péptidos, proteínas, ácidos nucleidos) tendo em conta que a carga destas substâncias depende do pH do meio em que se encontram. Moléculas negativa e positivamente mover-se-ão em direcções opostas do campo eléctrico.
A composição das proteínas pode ser determinada pelo processo da electroforese, comparando-se depois os patrões electroforéticos para as mesmas proteínas em diferentes espécies.
Toda uma variedade de moléculas pode ser individualizada por meio desta técnica.
Electroforese
Migração diferencial de compostos com carga quando sujeitos a um campo eléctrico.
A velocidade de migração é constante e depende do equilíbrio de forças e ao retardamento por atrito entre a amostra e o meio circundante.
A corrente é mantida do circuito pela electrólise que ocorre nos eléctrodos.
A velocidade da partícula num campo eléctrico calcula-se pela fórmula:
|
|
|
|
|
Z - nº de cargas de unidade e |
|
v |
= |
Z e E |
|
E - campo eléctrico |
|
f |
|
v - velocidade da partícula |
||
|
|
|
|
|
f - coeficiente de atrito |
Define-se mobilidade electroforética (u) como a velocidade de migração por unidade de campo eléctrico, numa mesma solução, partículas com a mesma carga podem ter mobilidade electroforética distinta pois podem possuir pesos moleculares diferentes: u=v/E, substituindo v temos que:
U = Ze/f em que o coeficiente de atrito (f) depende de do tamanho e forma da molécula e da viscosidade do meio.
Diferentes tipos de electroforese
Electroforese livre (frente móvel)Método introduzido por Tiselius em 1937.
As substâncias a separar introduzem-se num tubo em U, dissolvidas num tampão de pH adequado. Estabelece-se um limite entre a dissolução proteica e o tampão electroforético. Ao aplicar-se a corrente a frente proteica desloca-se por acção do campo. O avanço da frente pode seguir-se mediante observação UV.
Na década de 1945-55 deu-se um grande desenvolvimento na metodologia electroforética com a introdução de suportes inertes no meio, com o objectivo de evitar correntes de convecção produzidas pelo aquecimento na electroforese livre. Também o suporte permite a aplicação da amostra em bandas finas, com o que se consegue separações mais nítidas.
2.1. Electroforese em papel
O papel de filtro foi o primeiro suporte utilizado nos métodos electroforéticos de zona. O papel empapa-se com a solução tampão e coloca-se de modo conveniente entre os eléctrodos. A amostra aplica-se no centro ou em qualquer das extremidades do papel, dependendo das substâncias a separar e do pH do tampão.
Quando se querem separar substâncias de baixo peso molecular, como aminoácidos ou peptídeos, a difusão produzida pode evitar-se aumentando a diferença de potencial entre os eléctrodos reduzindo o tempo de separação. Este tipo de electroforese chama-se electroforese de alta voltagem.
A electroforese em papel pode também utilizar-se para o cálculo de pontos isoeléctricos.
O papel apresenta os inconvenientes de uma grande evaporação, maior na electroforese de alta voltagem, pelo que é necessário refrigerar o dispositivo de separação. Outro inconveniente é o fluxo electroendosmótico, movimento de arraste originado pelos iões do tampão de separação.
2.2. Electroforese em acetado de celulose
As películas de acetado de celulose trazem sobre o papel as vantagens de ser quimicamente mais homogéneo e tornar-se transparente, com o que as substâncias a separar, uma vez identificadas, podem quantificar-se mediante densitometria. Utiliza-se hoje com grande relevo nos laboratórios de análises químicas devido á sua fácil manipulação e rapidez.
2.3. Electroforese em gel
A utilização dos géis de amido e poliacrilamida na electroforese de zona foi um grande avanço devido a que com estes meios o gel não é um mero suporte que evita a difusão como também variando as concentrações do monómero, podem conseguir-se polímeros com crivagens diferentes. Desta forma a separação não se produz só por diferença de carga mas também por diferença de tamanho.
Shapiro, Vinuela e Maizel descreveram a electroforese em gel de poliacrilamida em presença do detergente aniónico dodecil sulfato de sódio (SDS) e indicaram que neste meio a separação é dependente do peso molecular. As proteínas em presença de SDS e beta-mercaptoetanol dissociam-se nas suas sub-unidades de modo que a separação electroforética permite o cálculo do peso molecular das cadeias polipeptídicas de proteínas oligoméricas. A mobilidade de uma cadeia polipeptídica pode ser só função do peso molecular se a carga por unidade de massa for constante ou se as propriedades hidrodinâmicas são unicamente função do comprimento da cadeia. Os dados que se possuem hoje sobre a acção do SDS nas proteínas indicam que o número de moléculas de SDS que se unem a uma cadeia polipeptídica é aproximadamente igual á metade do número de aminoácidos da cadeia, o que faz com que a carga por unidade de massa seja aproximadamente constante para todas as cadeias polipeptídicas. Por outro lado os estudos hidrodinâmicos e ópticos indicam que os complexos proteína-SDS se comportam como partículas alargadas nas quais o comprimento da partícula varia unicamente com o peso molecular da cadeia polipeptídica.
Hoje em dia a electroforese em gel na presença do SDS é um método muito utilizado para o cálculo dos pesos moleculares das proteínas oligoméricas. Quando se representa a mobilidade electroforética frente ao logaritmo dos pesos moleculares conhecidos de diversas cadeias polipeptidicas obtém-se uma recta que pode utilizar-se como padrão para o cálculo do peso molecular das sub-unidades da proteína que queremos estudar. Mediante esta técnica podem determinar-se pesos moleculares com uma precisão de mais ou menos 10 por 100.
Exemplo : Cálculo molecular da desidrogenase láctea u=0,455
|
Proteína |
Peso molecular |
Mobilidade |
|
Albumina do soro |
68000 |
0,18 |
|
Cataláse |
60000 |
0,22 |
|
Glutanato desidrogenáse |
53000 |
0,29 |
|
3-P-gliceraldeído desidrogenase |
36000 |
0,40 |
|
Tripsina |
23300 |
0,60 |
|
Mioglobina |
17200 |
0,74 |
2.4. Electroforese de disco
O meio para esta electroforese é a poliacrilamida. O método original de Ornstein e Davis (1964) estabelece duas descontinuidades de pH e uma descontinuidade de reticulação do meio electroforético.
A electroforese de disco pode distinguir famílias de proteínas, isómeros de carga ou isómeros de tamanho. Quando se representa o logaritmo da mobilidade relativa da proteína (distância percorrida pela proteína dividida pela distância percorrida por um corante que marque a frente electroforética), os isómeros de tamanho dão uma série de linhas não paralelas extrapoladas a um ponto comum próximo da concentração de acrilamida e as proteínas isómeros de carga dão uma família de linhas paralelas. As proteínas de diferentes cargas e tamanhos dão linhas não paralelas.
Aplicações práticas da electroforese
A engenharia genética depende da localização e da análise dos genes nos cromossomas e, em última instância, da sequenciação do ADN. As primeiras cartografias dos genes usavam os princípios da genética mendeliana. Assume-se que os alelos que são transmitidos em conjunto se localizam lado a lado num mesmo cromossoma: diz-se que estão ligados ou em linkage. Estes genes formam um grupo de ligação - grupo de linkage: passam para os mesmos gâmetas e são normalmente transmitidos em conjunto, de modo que não apresentam distribuição independente. Crossing-over que ocorra durante a meiose pode fazer com que estes alelos possam ser trocados entre os cromossomas de um par homólogo.
Pela análise das frequências de crossovers para muitos alelos diferentes pode traçar-se um mapa linear de cada cromossoma.
As técnicas modernas, mais do que o mapa dos genes, traçam o mapa do próprio ADN no interior dos genes: as posições de curtas sequências "marcadoras" são usadas como marcos de sinalização ao longo dos cromosssomas. Uma vez que um gene é descoberto, é necessário desvendar a sua sequência de bases antes de a sua função ser estudada. A sequenciação tornou-se mais fácil com o desenvolvimento de métodos de clonar o ADN - fabricar grandes quantidades de fragmentos idênticos. No método de sequênciação do ADN mais usado, a cadeia é desnaturada em cadeias simples. Estas são então utilizadas como moldes para a síntese do ADN, mas de tal modo que a replicação pára quanto a dupla hélice em crescimento atinge uma determinada base do molde. Para além de se fornecer polimerase do ADN e as quatro bases, A - G -C -T, também se usam pequenas quantidades de didesoxinucleótidos destas bases. Esta incorporam-se, como as bases normais, na dupla hélice em crescimento, mas impedem a continuação da cadeia. Os fragmentos são então separados por electroforese em gel e a sequência de bases do segmento de ADN original pode ser facilmente deduzida.
Se os fragmentos de ADN forem radioactivamente marcados, as bandas correspondentes a fragmentos de tamanhos diferentes podem ser detectadas colocando o gel de electroforese sobre uma película fotográfica. Esta técnica está a ser usada para sequenciar o genoma humano, que contém centenas de milhares de pares nucleótidicos. É muito mais rápida que a análise de proteínas e, por essa razão, as estruturas de muitas proteínas são estudadas a partir da análise dos fragmentos de ADN que as codificam, deduzindo depois as sequências de aminoácidos. O genoma de vários organismos "modelo" está a ser cartografado tendo em vista proporcionar o material de partida para se estudar a função dos genes.
|
|
|
|
|
|

A engenharia genética também tem um papel a desempenhar na prevenção de doenças não hereditárias. As vacinas podem passar a ser vírus modificados, inofensivos, como o vírus Vaccinia, que está a ser modificado para a preparação de vacinas contra a raiva, o herpes e a gripe. Pode inserir-se um máximo de vinte cinco genes extra no genoma viral para se obter imunidade múltipla. Outras vacinas, como a da hepatite B, podem simplesmente ser as proteínas superficiais do revestimento do vírus em questão. O gene viral é inserido num microorganismo, que produz depois a vacina.
A dactiloscopia genética é uma técnica utilizada na ciência forense onde se analisam as diferenças entre segmentos de ADN chamados ADN minissatélites. Este ADN encontra-se entre os genes e é utilizado na dactiloscopia porque é extremamente viável. A hipótese de dois indivíduos sem qualquer relação de parentesco possuírem exactamente as mesmas regiões minissatélites é muito remota, por isso a dactiloscopia do ADN constitui um teste viável. A região minissatélite é cortada em fragmentos por meio de enzimas de restrição. Os fragmentos daí resultantes são separados de acordo com o tamanho, por uma técnica chamada de electroforese em gel, em que os fragmentos são impelidos por um campo eléctrico através de um gel.. Isto produz uma "escada" de ADN, em que cada "degrau" corresponde a um fragmento de tamanho diferente. As escadas produzidas pelo ADN de indivíduos diferentes podem ser comparadas ; as diferenças são facilmente detectadas e as pessoas distinguidas.
|
|
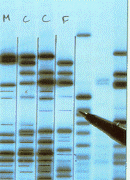
As bandas obtidas por dactiloscopia que se encontram representadas estão marcadas com um M de Mãe, um C de Criança e um F de Pai (em inglês, father). Ambas as crianças partilham algumas bandas com cada progenitor, o que prova que o indivíduo de que foram extraídas as amostras são, de facto aparentados. Uma falta de correspondência indica a ausência de parentesco.
Esta técnica rotineiramente utilizada em casos de paternidade contestada pode ter também algumas aplicações menos vulgares, em1992 foi utilizada para confirmar que cinco esqueletos exumados de uma campa nas imediações de Ekaterinburg eram os do Czar Nicolau II da Rússia, da sua mulher e das três filhas.
As técnicas sofisticadas da medicina e da prática forense permitem que a compreensão crescente do ADN seja aplicada com resultados práticos. Por exemplo, generalizaram-se os testes de ADN para identificar e prever doenças genéticas. Pelo estudo da árvore genealógica das famílias afectadas é possível traçar um quadro das relações genéticas e prever quais os indivíduos em risco que podem depois ser sujeitos a despistagem, em busca de deficiências genéticas. Uma técnica bastante comum recorre a certas enzimas, chamadas endonucleases de restrição, para cortar o ADN em determinados pontos. Os fragmentos são então separados e identificados por sondas de ADN. A este processo dá-se o nome de análise dos RFLP (restriction fragment length polimorfism).
Vários métodos de despistagem podem ser usados para analisar um feto em desenvolvimento em busca de afecções hereditárias como o sindroma de Down. A amniocentese, a biópsia das vilosidades coriónicas e a cordocentese são técnicas pelas quais as células ou as proteínas fetais podem ser obtidas, postas em cultura e usadas para diagnóstico genético.
A genética forense tem outras aplicações. As "impressões digitais" do ADN representam uma valiosa ferramenta para a polícia científica. À semelhança do que acontece com uma impressão digital vulgar, a impressão genética é exclusiva de cada indivíduo (com excepção dos gémeos verdadeiros). A sua determinação implica a observação de sequências específicas de ADN, que pode ser obtido de amostras extremamente pequenas de tecidos, cabelo ou sangue, eventualmente deixados no local do crime. Apenas cinquenta microlitros de sangue, cinco microlitros de sémen ou dez raízes de cabelos são o suficiente, assim como secreções bocais e células do feto. Para além da sua utilização na captura de criminosos, em especial os violadores, as impressões genéticas podem ser usadas para estabelecer relações familiares. As pessoas envolvidas na conservação das espécies usam-nas para terem a certeza de que a procriação em cativeiro se faz entre indivíduos que não pertencem à mesma família.
|
Junho, 2000 |
Música de Fernando de Brito Vintém |